




Chiba Medical J. 101E:39-48, 2025
doi:10.20776/S03035476-101E-3-P39
〔 Original Article 〕
Maki Fujita1), Hirokazu Usui1), Saori Nakahashi1,2)
Hiroshi Ishikawa1), Tatsuya Kobayashi1,3), Naoki Aoyama4)
Yasunori Sato5), Yasuhito Michikura6), Keiichi Kato4)
and Makio Shozu1,7)
1) Department of Reproductive Medicine, Obstetrics and Gynecology, Graduate School of Medicine, Chiba University, Chiba 260-8670.
2) Department of Nephrology and Hypertension, Blood, Saitama Medical Center, Blood purification, Saitama 350-8550.
3) Department of Regulatory Science, Fujita Health University, Tokyo 144-0041.
4) Kato Ladies Clinic, Tokyo 160-0023.
5) Department of Preventive Medicine and Public Health, Keio University School of Medicine, Tokyo 160-8582.
6) Kanazawa Tamago Clinic, Ishikawa 920-0994.
7) Evolution and Reproduction, Medical Mycology Research Center, Chiba University, Chiba 260-8673.
(Received February 22, 2025, Accepted May 19, 2025, Published September 10, 2025.)
【Objective】Tripronuclear (3PN) zygotes derived from conventional in vitro fertilization (c-IVF) exhibit diandric triploidy due to dispermic fertilization. These zygotes may spontaneously convert to diploid and develop into morphologically normal blastocysts, potentially making them candidates for embryo transfer. However, limited evidence confirms that these diploid blastocysts have biparental inheritance. This study aimed to determine the parental origin of chromosomes in 3PN zygotes and assess their potential conversion to biparental diploidy.
【Methods】Tripronuclear zygotes observed in c-IVF culture were analyzed. After the first mitotic division, day 2 embryos were dissociated into single blastomeres (2?5 cells per embryo), lysed, and subjected to whole-genome amplification using Phi 29 DNA polymerase, followed by short tandem repeat polymorphism analysis. Ploidy determination was performed via 25?72 PCR reactions per zygote, using an aliquot of the diluted lysate as the PCR template.
【Results】A total of 509 PCR reactions were conducted for 11 3PN zygotes, yielding 84 amplicons in 82 reactions: a single amplicon in 80 (15.7%) reactions and two amplicons in four (0.4%) reactions. The remaining 427 (83.9%) reactions produced no amplicons. The parental origin of chromosomes was estimated as follows: biparental diploid origin (one paternal and one maternal allele each) in 4 zygotes (36%), diandric triploid origin (two paternal alleles and one maternal allele) in 6 zygotes (55%), and androgenetic origin (two paternal alleles and no maternal allele) in 1 zygote (9%).
【Conclusions】These findings suggest that c-IVF-derived 3PN embryos (triploid) may convert to biparental diploid during the first mitotic division. However, further studies using comprehensive and complementary methods are needed to confirm their suitability for embryo transfer.
pronucleus, in-vitro fertilization, triploid, short repeat polymorphism
Tripronuclear (3PN) zygotes are one of the most frequent chromosomal abnormalities observed in human conventional in vitro fertilization (c-IVF), occurring in 1%?4% of c-IVF zygotes[1]. Researchers have analyzed the karyotype of these zygotes and found that most 3PN zygotes after c-IVF are diandric monogynic triploids (two paternal chromosome sets and one maternal chromosome set)[2-4]. This suggests that these zygotes are fertilized by two sperm.
Tripronuclear zygotes have been considered inappropriate for embryo transfer and are usually discarded. However, they may develop into normal-appearing blastocysts[5]. They are not used for embryo transfer due to concerns about chromosomal abnormalities[2,3,6-8]. Karyotyping of these blastocysts has confirmed that they primarily consist of triploid cells of dispermic origin[9,10].
However, researchers have also found that 21?35% of 3PN-zygote-derived embryos are composed of diploid cells, suggesting that those embryos self-convert from triploid to diploid before reaching the blastocyst stage[2,6,7,11]. Pang et al. examined the ploidy of 3PN-derived embryos using FISH analyses and found that some 3PN-derived 2?4-cell stage embryos were diploid[11]. They concluded that re-diploidization occurs as early as the 2?4-cell stage. These results support the idea that some 3PN embryos self-convert to diploid cells. However, the biparental origin of these rediploidized blastomeres in early cleavage-stage embryos remains unclear.
To address this gap, we developed a single-cell PCR-based STR analysis and examined the DNA origin of individual blastomeres from 2?4-cell stage embryos.
IVF-ET and 3PN zygotes
Tripronuclear zygotes (n = 12) were produced from c-IVF cycles conducted for infertile couples (n = 12) due to unexplained infertility or tubal factors. Oocytes were retrieved after treatment with human menopausal gonadotropins (n = 3), clomiphene (n = 3), clomiphene plus human menopausal gonadotrophin (n = 5), or no medication (n = 1). Thirty-two to 34 hours after the triggering maturation using buserelin (100 μg, nasal) or human chorionic gonadotropin (10,000 IU), oocytes were retrieved. The oocytes were cultured individually in 30 μL of human tubal fluid medium (Irvine Scientific, Santa Ana, CA, USA) supplemented with 10% Serum Substitute Supplement™ (Irvine Scientific) under mineral oil at 37℃ in 5% CO2, 5% O2, and 90% N2 for 3 h. Metaphase II oocytes were inseminated with sperm prepared using the ISolate® Kit (Irvine Scientific) to achieve a final concentration of 1 x 106 sperm/mL. Eighteen hours after insemination, zygotes were observed under a phase-contrast microscope. Zygotes showing three pronuclei, roughly in equal size, and two polar bodies, which should be discarded from the treatment protocol, were collected for this study. Tripronuclear zygotes with two equal-sized pronuclei associated with an extremely small surplus pronucleus less than 15 μm in diameter were excluded from this study[12]. The zygote, so called 2.1PN embryo, were not included in this study, because they are essentially considered to be diploid[13].
Isolation of blastomeres
Tripronuclear zygotes were cultured further until the first cleavage was observed, usually by 8?22 h post-fertilization (Fig. 1). Then, 2?5-cell stage embryos were cryopreserved using the Cryotop Safety Kit for vitrifying oocytes and embryos (Kitazato BioPharma, Fuji City, Shizuoka, Japan) and preserved in liquid nitrogen. Cryopreserved embryos were thawed using the Cryotop Safety Kit-Thaw (Kitazato BioPharma) to isolate blastomeres. Embryo viability was verified after a 2-hour cultivation in complete Early Cleavage Medium (Irvine Scientific). The viable embryos were treated with acidic Tyrode’s solution (138 mM NaCl, 2.7 mM KCl, 1.6 mM CaCl2・2H2O, 0.4 mM MgCl2・6H2O, 5.56 mM glucose, and 0.1 mM polyvinylpyrrolidone) to remove the zona pellucida. After washing three times with calcium/magnesium-free phosphate-buffered saline (PBS), the zona pellucida-free embryos were transferred to Hank’s balanced salt solution (Life Technologies, Carlsbad, CA) and separated into individual blastomeres by mouth-controlled pipetting. The resulting single blastomeres were washed three times in 1% polyvinylpyrrolidone in PBS droplets covered by mineral oil. Each blastomere was recovered into a 200-μL collection microtube containing 1.0 μL of distilled water under a stereo-microscope. Microtubes containing single blastomeres were centrifuged and stored at -80℃ for 1 week or less until used in the experiment.
STR polymorphism analysis of parental genome
We performed STR polymorphism analysis and selected informative loci to differentiate the maternal and paternal origins of the genomes.
Genomic DNA was extracted from peripheral blood or buccal swabs of pairs using the QIAamp DNA Blood Mini kit (Qiagen GmbH, Hilden, Germany) and subjected to multiplex STR polymerase chain reaction (PCR) analysis using the PowerPlex 16 multiplex STR system (Promega Corporation, Madison, WI, USA) [14]. Amplicons were dissolved using the ABI PRISMR 310 Genetic Analyzer (Applied Biosystems, Inc., Foster City, CA). Data was analyzed using Gene Mapper software 4.0 (Applied Biosystems, Inc.). We selected one informative locus per embryo, where the two paternal alleles had different numbers of repeats (heterozygous) and were also distinct from the maternal allele (s). The STR analysis allowed us to differentiate biparental triploid from biparental diploid zygotes.
Single-cell PCR for STR analysis
Whole genome amplification
The isolated blastomere in microtubes containing 1.0 μL of water was thawed and digested with 1.0 μL of proteinase K (1 μg/mL) and 0.4 μL of proteinase K buffer (3.0% sodium dodecyl sulfate, 0.06 M Tris-HCl, 0.03 M ethylenediaminetetraacetic acid) for 30 min at 50℃. Proteinase K was inactivated by incubating at 80℃ for 10 min.
The genomic DNA was amplified using the GenomiPhi version 2 DNA amplification kit (GE Healthcare, Tokyo, Japan) according to the manufacturer’s instructions with the following modifications. Nine μL of sample buffer supplied with the kit were added directly to the lysate, incubated at 97℃ for 3 min, and then quickly chilled on ice. Then, 9.0 μL of the reaction buffer and 1.0 μL of enzyme mix containing Phi 29 DNA polymerase diluted 100-fold with 50 mM Tris-HCl were added to the reaction tube, which was incubated for 3 h at 30℃ and then heated to 65℃ for 10 min for enzyme inactivation.
The amplified DNA was purified using the MinElute Reaction Cleanup Kit (Qiagen Sciences, Germantown, MD). The DNA was eluted by applying 16?18 μL of Tris-EDTA solution to the column, yielding 12?16 μL of eluate.
STR-PCR of whole genome amplification samples
The informative locus was PCR-amplified from whole-genome amplified (WGA) DNA using in-house designed primer pairs (Table 1).
Each PCR reaction consisted of 1 μL of WGA eluate and 4.0 μL of the reaction mixture, containing 0.5 μM primer pairs, 1 × LightCycler FastStart DNA Master SYBR Green (Roche Diagnostics, Basel, Switzerland), 3 mM MgCl2, and 0.0625 μL of LightCycler Uracil-DNA glycosylase (Roche Diagnostics). Real-time PCR was performed using the LightCycler 1.5 instrument (Roche Diagnostics).
The cycling program was as follows: 10 min at 40℃ to activate uracil-DNA glycosylase, 10 min at 95℃ to activate Taq polymerase, followed by 45 amplification cycles of 5 s at 95℃, 10 s at the annealing temperature of each primer (Table 1), and 10 s at 72℃. The temperature was then gradually raised from 65℃ to 95℃ for melting, followed by 30 s at 40℃ for cooling.
The PCR products were separated by electrophoresis in a 12% polyacrylamide gel using 1 × Tris/borate/ EDTA buffer. Eleven to fourteen PCR reactions were performed collaterally for each blastomere, resulting in a total of 25-72 reactions per embryo (Fig. 1).
Statistical analysis
Mann-Whitney test was used to compare paternal/ maternal allele ratios. A probability < 0.05 was considered significant.
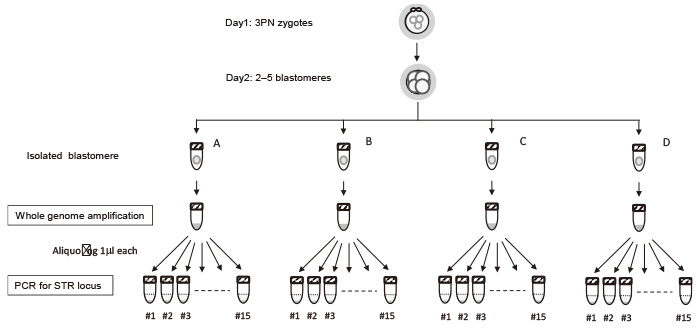
Fig. 1 Sample preparation flow diagram for short tandem repeat PCR. Each blastomere lysate was aliquoted for 11-15 PCRs, resulting in 25-72 PCRs per zygote.
Table 1 Primer sequences and annealing temperatures

Single-cell PCR-based ploidy analysis
All 3PN-derived zygotes (n = 12) were viable and cleaved to 2?5 blastomeres on day 2 of in vitro culture. One zygote was lost during preparation, and 38 blastomeres from the remaining 11 3PN-derived embryos were analyzed (Table 2).
A total of 509 PCR reactions yielded 84 amplicons in 82 reactions: one amplicon in 80 (15.7%) reactions and 2 (0.4%) amplicons in two reactions (Fig. 2). The remaining 427 (83.9%) reactions had no amplicon. This right-skewed distribution was in good agreement with the theoretical distribution calculated with a probability of 0.044 for the locus DNA inclusion probability, probability that any one of allele detected by single PCR run) (Fig. 3 and Supplementary materials). This low probability suggested that most amplicons detected by the PCR were from a single copy of allele DNA. Out of 38 blastomeres, 32 blastomeres yielded ? 1 amplicon (s), while 6 blastomeres yielded no amplicons.
Ploidy and origin of genomic DNA
Allelic status was determined in 10 of 11 zygotes: Six were diandric monogyny triploid, and four were biparental diploid (Table 2). The remaining one had seven amplicons from the paternal long allele and none from the maternal allele, which was estimated to be diandric diploid.
Representative cases
Representative cases are shown in Figure 4. A total of 25 PCR reactions for the D13S317 locus was performed for WGA-derived DNA from three blastomeres (embryo #10), resulting in six amplicons: three from a long paternal allele and three from a long maternal allele (Fig. 4A), suggesting that the embryo is biparental diploid. In embryo #1, 37 PCR reactions (D8S1179 locus) for three blastomeres yielded six positive amplicons: two from a short paternal allele, three from a long paternal allele, and one from maternal allele (Fig. 4B). This suggests that the embryo is diandric monogynic triploid and is heterozygous for androgenetic alleles.
Paternal to maternal allele amplicon ratio
As mentioned above, each amplicon is virtually considered to be derived from a single genome set; therefore, the ratio between amplicon types reflects the ratio of the source chromosome set.
We calculated the ratios between paternal and maternal amplicons for each zygote (Fig. 5). In three of the estimated diploid embryos, the ratios were 1.0 or lower, whereas in all six triploid embryos, the ratios were 2.0 or higher. This difference was statistically significant (p = 0.014).
Table 2 Summary of STR-PCR results of blastomeres and estimated genetic origin
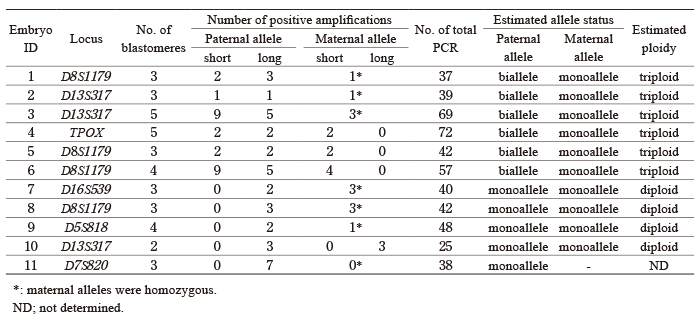
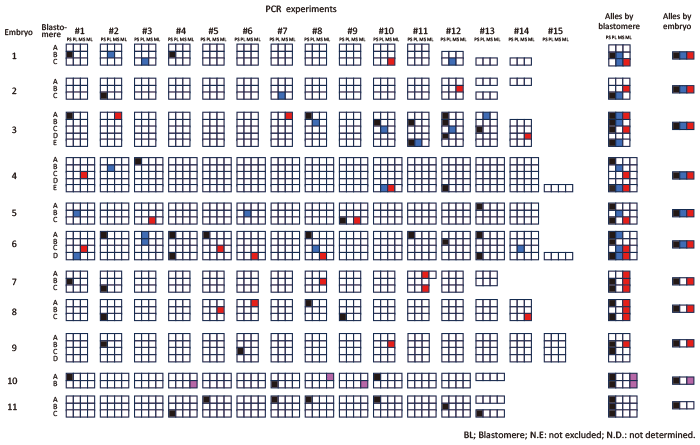
Fig. 2 Visualization of allelic profiles by PCR run and embryo. A set of three or four cubes arranged in a row represents the result of one PCR run for a single blastomere. A?E denote the blastomeres of each zygote. The numbers in the first row (#1?#15) indicate groups of PCR runs conducted on individual blastomeres derived from the same WGA DNA. Blank cubes indicate negative PCR results. Alleles detected in each PCR experiment are represented by colored cubes: black indicates the short paternal allele (PS), blue indicates the long paternal allele (PL), red indicates the short maternal allele (MS), and pink indicates the long maternal allele (ML) . If the maternal allele is monovalent, the detected allele is shown in red. The results are summarized by blastomere and by embryo in the second column (alleles by blastomere) and the third column (alleles by embryo), respectively.
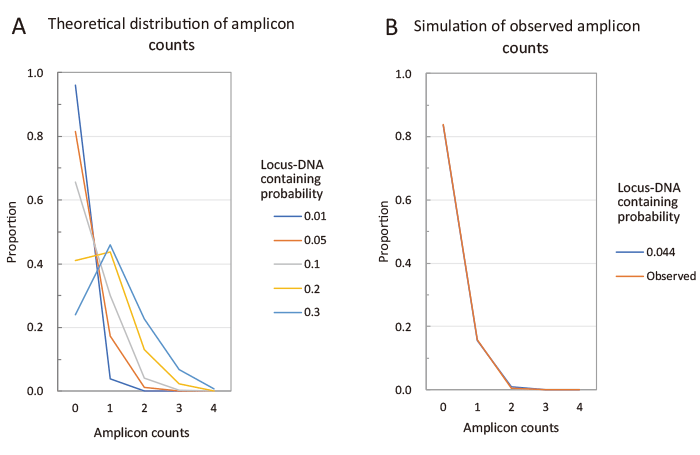
Fig. 3 Simulation of the distribution of the number of amplicons detected in each PCR run. Each PCR run can detect 0?4 different amplicons. The number of amplicons detected per run (amplicon count) depends on the amount of template DNA present in the PCR tube. We calculated the proportion of each amplicon count based on the locus DNA containing probability that 1 μL of whole genome amplification (WGA) eluate used as a PCR template contains one copy of the target locus (denote as C). Detailed calculations are provided in the supplementary materials. Figure A shows the simulated distribution of amplicon counts. As C increases from 0.01 to 0.3, the distribution shifts from a right-skewed shape to one with a peak near 1. Figure B shows the distribution observed in this study. This observed distribution closely matches the simulated distribution with C = 0.044, for which the residual sum of squares is less than 0.0001 (Supplementary materials). To simplify the calculation, the following assumptions were made: (1) Each embryo has four different alleles. (2) PCR amplification from a single copy of DNA always succeeds.
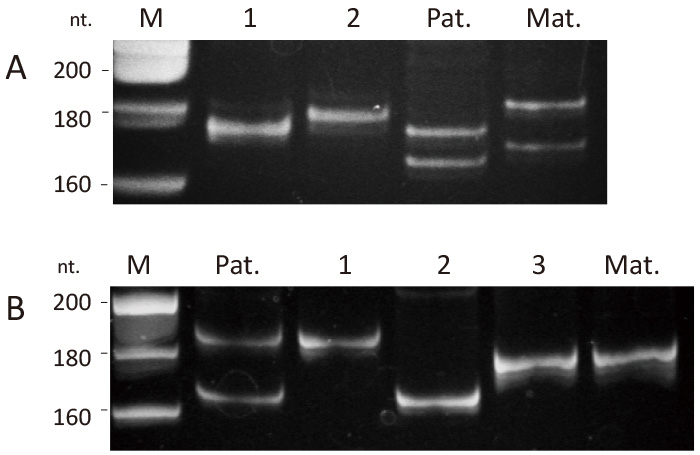
Fig. 4 Representative Polyacrylamide Gel Electrophoresis. (A) Two types of D13S317 amplicons were detected for embryo #10, corresponding to the long paternal allele (lane 1) and the long maternal allele (lane 2). PCR reactions with no amplicons are not included in this electrophoresis. (B) Three types of D8S1179 amplicons were detected for embryo #1: two corresponding to the long and short paternal alleles (lanes 1 and 2) and one corresponding to the maternal allele (lane 3). M, DNA size marker; Pat, paternal blood; Mat, maternal blood.
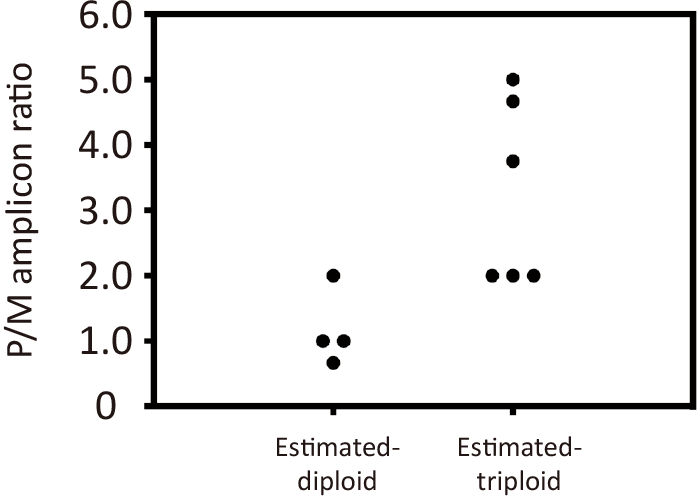
Fig. 5 Paternal to maternal allele amplicon ratio. The total number of paternal amplicons detected in the PCRs was divided by the number of maternal amplicons to calculate each embryo’s paternal/maternal amplicon ratio. The ratio between the diandric monogynic triploid and biparental-diploid estimated groups was compared.
The present study demonstrated that nearly half of the 3PN-derived early-stage embryos were diandric triploid, while the other half were biparental diploid. Given that most 3PN zygotes observed in c-IVF are triploid due to fertilization by two sperm, our findings suggest that some 3PN zygotes may spontaneously convert from triploid to diploid during the first cleavage through the selective elimination of a sperm-derived chromosome set.
Diploidization can produce a biparental embryo
The present study focused on the origin of chromosome sets in 3PN-derived embryos. In c-IVF, most 3PN zygotes result from dispermic fertilization and are thus diandric monogynic triploid[7]. The results of the present study suggest that diploidization can occur by loss of either the paternal (#7?10) or maternal chromosome set (#11), but the loss of the paternal chromosome set is more likely to occur.
The loss of one paternal chromosome set gives rise to biparental diploid embryos, which may be suitable for embryo transfer. In contrast, losing the maternal chromosome set results in diandric diploid embryos, which may develop into a complete hydatidiform mole[15]. The loss of the paternal chromosome set occurred four times more frequently than the loss of the maternal chromosome set, suggesting a bias toward a pattern favorable for embryonic development. More extensive studies are necessary to determine whether diploidization more frequently produces biparental embryos than diandric embryos.
Possible mechanism of diploidization
Several mechanisms may explain the diploidization during the early stage of development. One is prematurely separating small cytoplasmic fragments containing supernumerary pronucleus through premature cytokinesis, resulting in a diploid zygote before the first cleavage[16]. Another mechanism is the altered cleavage of zygote[6]. Tripronuclear zygotes often exhibit altered cleavage patterns, such as direct cleavage to 3-cells at the first mitotic cleavage. Tripronuclear zygotes may form tripolar spindle apparatus and directly cleave to 3-cells, which may result in restoration of diploidy during the first mitosis [6,17]. In line with this, we observed that 10 of 11 zygotes cleaved to 3 or more cells on day 2, four of which diploidized. Therefore, altered cleavage may be involved in diploidization, although complementary studies with larger sample sizes are needed to prove this. It should be noted, however, that such modified division may be associated with complex chromosomal anomalies, including multiple aneuploids.
Single-cell STR for estimation of parental origin
We developed the single-cell STR assay to determine the origin of chromosome sets. The sensitivity and accuracy of this assay depend on the efficacy of whole genome amplification. The estimated locus DNA containing probability was as low as 0.044, suggesting that the efficacy of the genomic amplification was less than expected. The kit that we used was the early model, and recent kits would improve the efficacy.
Next-generation sequencing (NGS) is now clinically used for preimplantation genetic analysis of aneuploidy (PGT-A). Moreover, NGS plus single nucleotide polymorphism analysis can identify the origin of chromosome sets. A recent study used NGS to 3PN-zygotes-derived blastocysts and found that 37.8% (14 of 37) were diploid but did not report on the parental origin of chromosomes[18]. A drawback of NGS-based analysis is that 5-10 cells are usually required for a reliable and accurate diagnosis. Obtaining 5-10 cells from day 2 embryos is virtually impossible. However, NGS studies using 1?2 cells from early cleavage stage embryos have been reported[19].
Chromosomal variability and developmental potential in 3PN zygotes
The potential for conversion from 3PN may depend on their chromosomal constitution, which is influenced by the mechanism of extra pronucleus formation. Several mechanisms contribute to 3PN formation, including: diandric triploidy, resulting from polyspermy; digynic triploidy, which arises from either the failure to extrude the second polar body or endoduplication of the female pronucleus; diploid or mosaic chromosomal patterns, associated with abnormal mitotic division, including aberrant spindle function; and 2.1PN oocytes, which contain a small part of diploid chromosome set as an accessory pronucleus[12,13,18,20-25]. Among these, polyspermy is considered the leading cause of 3PN formation in c-IVF, although other mechanisms can also contribute[1].
Our results indicate that at least six 3PN zygotes were of dispermic origin. These zygotes exhibited blastomeres with different allele combinations, suggesting a complex process of karyokinesis.
In contrast, three zygotes contained three blastomeres each, which were considered biparental diploid. A dispermy-derived 3PN zygote undergoing three-cell cleavage in the first mitotic division could, in theory, produce biparental diploid blastomeres, each with a different paternal allele. However, no such embryo was observed in our study. To confirm whether these diploid blastomeres originate from true triploid 3PN cells, further research with a larger sample size is necessary.
Clinical relevance of this study
There are several applications for 3PN-derived diploidized embryos. First, self-diploidized embryos have been used for embryo transfer in IVF treatment [26]. Embryos confirmed to have a normal karyotype by PGT-A may be considered for transfer[27]. However, one report suggests routine screening of 3PN-derived embryos after ICSI is not recommended due to cost-effectiveness concerns [5].
Second, surplus pronuclei have been surgically removed using micromanipulators, and the resulting diploid embryos have been used for embryo transfer or the establishment of embryonic stem cells[28]. Live births have been reported following the removal of the extra male pronucleus[29]. Additionally, the removal of surplus pronuclei has been shown to enhance the establishment of human embryonic stem cell lines [30]. These methods provide an alternative resource for human cell studies.
The small sample size is a limitation of this study. Another limitation is that only one STR locus was examined. Due to this limitation, we could not consider the possible mosaicism of embryos in our discussion. Possible allele dropout is another limitation[31]. During whole-genome amplification, locus-specific amplification errors may occur, leading to misreading allele numbers. Although we cannot exclude this possibility, we do not believe that it happened frequently in our study, as maternal alleles were successfully detected in all blastomeres except one, which may have been a hydatidiform mole with no maternal chromosome set.
We have provided evidence that c-IVF-derived 3PN zygotes may give rise to diploid embryos of biparental origin. However, further studies are necessary to determine whether these embryos have normal chromosomes suitable for the transfer.
M. F.: conception and design, acquisition of data, data analysis, and drafting of the article. H. U.: acquisition of data, data analysis, manuscript revision, and final approval.: acquisition of data, drafting the article, and final approval. S. N., T. K. and H. I.: sample preparation, final approval. N. A., Y. M., and K. K.: patient recruitment, clinical information, sample correction, conception, and design. Y. S.: analysis and interpretation of data. M. S.: conception and design, data analysis and interpretation, manuscript revision, and final approval.
This work was supported by research grants from Chiba University (M. F.) and Grant-in-Aid for Scientific Research (C) (no. 23592435 to H. U.), Grant-in-Aid for Scientific Research on Innovative Areas (no. 23132503 to M. S.) from the Ministry of Education, Culture, Sports, Science and Technology of Japan, and Health and Labour Sciences Research Grants for Research on rare and intractable diseases (H23-090 to M. S.) from the Ministry of Health, Labour, and Welfare.
M. S. is a member of the Editorial Board of the Chiba Medical Journal but was not involved in the review or acceptance of this paper.
This study was approved by the Institutional Review Board of Chiba University Graduate School of Medicine (#284), and written informed consent was obtained from all patients and their partners before enrollment.
The data of this study are available from M. F. upon reasonable request.
We thank Kato Ladies Clinic, Kanazawa Tamago Clinic, and Towako Ladies Clinic members for their kind assistance in sample collection.
Address correspondence to Dr. Makio Shozu.
Evolution and Reproduction, Medical Mycology Research Center,
Chiba University, 1-8-1 Inohana, Chuo-ku, Chiba 260-8673,
Japan.
Phone: +81-43-222-7171.
Fax: +81-43-226-2486.
E-mail: shozumakio@chiba-u.jp
